What are Viroids? Understanding the World’s Smallest Pathogen
(Posted on Wednesday, February 22, 2023)
Viruses, bacteria, and fungi; most people will have heard of these three major types of disease-causing microbes. Some will also have heard about protozoa, which are larger and more complex than the aforementioned three. The particularly studious may even know of prions, which are “infectious”, mis-folded proteins that cause neurodegenerative diseases, including mad cow disease. But few will know about the smallest infectious pathogens discovered to date: viroids.
These biological oddities are to blame for a host of different crop diseases, causing huge losses across many different agricultural industries. In humans, the viroid-like hepatitis D virus (HDV) can cause joint pain, inflammation of the liver, and extreme fatigue. Although people can overcome the acute phase of infection, for many the disease becomes chronic. Coinfection of hepatitis B and D is considered the most severe form of hepatitis, with patients frequently developing serious complications ranging from extreme fatigue to full-scale liver cirrhosis.
So, what are viroids and how do they replicate? What follows is a brief overview of what we know so far.
Nature’s Minimalist
Every pathogen carries with it a toolkit for infection — the types of tools in the kit, and the size of the kit, however, can vary significantly.
Viruses generally travel quite light. They take only what they need and depend on the host cell for the rest, including the biological machinery that allows them to make copies of themselves. Hijacking the host cell translation system permits viruses to produce the proteins encoded in their genetic material. Some of these proteins will be used to further copy the viral genome, enabling effective replication — every virus encodes at least one protein to help with this process. Others give a virus its structure and protective coating. These are necessary for viral entry into host cells. And in some cases, a virus may also carry proteins that help give it a competitive edge over our immune system, letting it go undetected or actively suppressing our defenses. Regardless, the function of the genetic material —whether in the form of DNA (deoxyribonucleic acid) or RNA (ribonucleic acid)— is to serve as a blueprint for the production of proteins. DNA and RNA do the instructing, proteins do the actual legwork.
Viroids take this lightweight approach to the extreme: they are small, covalently closed circular (ccc) RNA molecules that, crucially, do not encode any proteins. Where a virus such as SARS-CoV-2 is made up of nearly 30,000 nucleotides —the basic building blocks of DNA and RNA— viroids only contain between 220 to 450 nucleotides. This is more than five times smaller than the smallest known virus. And since viroids lack a protein “coat” they are completely naked, nothing but RNA.
The minimalism of viroids does come with the drawback of being entirely dependent on passive mechanisms of transmission. Since they lack a protein coat, they do not have any surface proteins and cannot recognize, bind to, and enter healthy cells. The only way for a viroid to get into a cell is if there is pre-existing damage to the cell membrane, affording it an opportunity for entry. Otherwise, it is stuck outside the door.
“Rolling-Circle” Replication and Ribozymes
If viruses rely on their proteins for replication, how is it that viroids manage without? They take advantage of a process called “rolling-circle replication” (Figure 1). This mechanism was discovered by my PhD mentor, Walter Gilbert, and fellow graduate student, David Dressler, in the 1970s. For the most part, rolling-circle replication serves to create duplicates of DNA, but viroids and some RNA viruses —neither of which possess any DNA— have managed to appropriate the task for their own benefit.
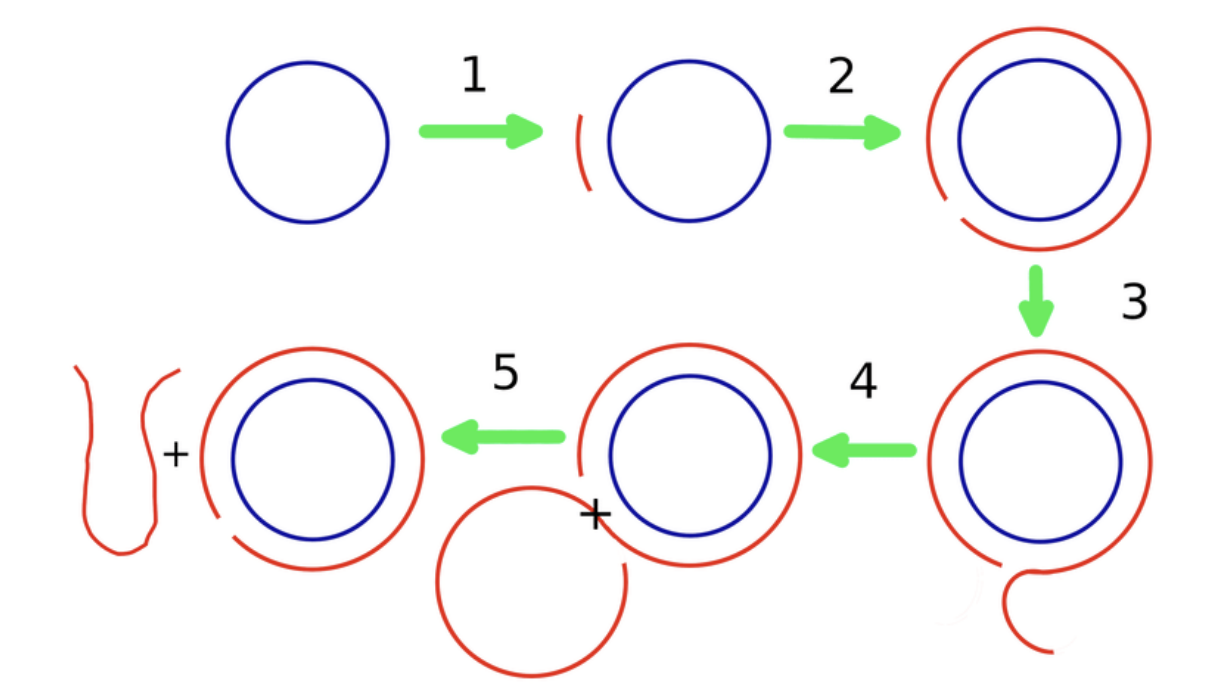
FIGURE 1. Schematic diagram of the rolling circle replication process. 1: A short complementary primer attaches to a circular ssRNA template. 2: It is extended by the template-directed primer extension process towards full length. 3: Upon attaining full length, the primer extends further by displacing the other end of itself from the initial point, resulting in an overhanging portion. 4: when the overhanging tail attains a length equal to that of the circular template, it breaks apart. 5: The separated tail becomes an open-ended ssRNA. SOURCE: “Evolution towards increasing complexity through functional diversification in a protocell model of the RNA world” ROY ET AL. 2021
First, the single-stranded viroid RNA is copied by a piece of host-cell machinery called RNA polymerase II. This is already unusual, since RNA polymerase II traditionally transcribes DNA into RNA, not RNA into RNA. Somehow, viroids manage to subvert this and hijack the enzyme. RNA polymerase II produces a long, fused chain made up of multiple copies of the viroid genome. But since the genomes are fused together, they need to be cleaved at the appropriate places before they are fully functional.
Cleavage of the long, multi-genome chains can happen in one of two ways, and viroids are classified into families depending on the method they use (Figure 2). The first family of viroids, called Pospiviroidae, are rod shaped. These rely on conserved genetic motifs in their RNA being recognized and cleaved by host enzymes — current consensus flags RNA polymerase III as the most likely candidate. Once the long chain of viroid genomes has been chopped up into single, free-floating pieces, another host enzyme, called DNA ligase I, fuses the ends of each strand together, shaping them back into their usual circular form. Again, DNA ligase I is actually meant for DNA replication, but the viroids manage to subvert the mechanism to suit their needs.

FIGURE 2. Asymmetric and symmetric variants of the rolling-circle mechanism proposed for replication of members of the families Pospiviroidae and Avsunviroidae, respectively. Orange and blue colors refer to plus and minus polarities, respectively, with cleavage sites denoted by arrowheads. The enzymes and ribozymes that presumably catalyze the replication steps are indicated. Notice that RNA polymerase II (and NEP) is redirected to transcribe RNA templates and DNA ligase 1 to circularize RNA substrates. Abbreviations: HHRz, hammerhead ribozyme; NEP, nuclear-encoded polymerase. SOURCE: “Viroids, survivors from the RNA world?” FLORES ET AL. 2014
The second family of viroids, called Avsunviroidae, does not depend on host-cell machinery for cleavage. Instead, these viroids make use of so-called “hammerhead” ribozymes (HHRz). Although this may sound like a protein, it is not; a ribozyme is a strand of RNA that is capable of acting like an enzyme, catalyzing biochemical reactions such as RNA splicing. Rather than simply serving as a set of genetic instructions for protein synthesis, RNA in the form of a ribozyme does the manual labor itself. So, viroids of the Avsunviroidae family carry with them the tools to self-cleave, which is integral to the replication process (Figure 3). Unlike their counterparts, Avsunviroidae have a branched, hammerhead-like shape (Figure 4).
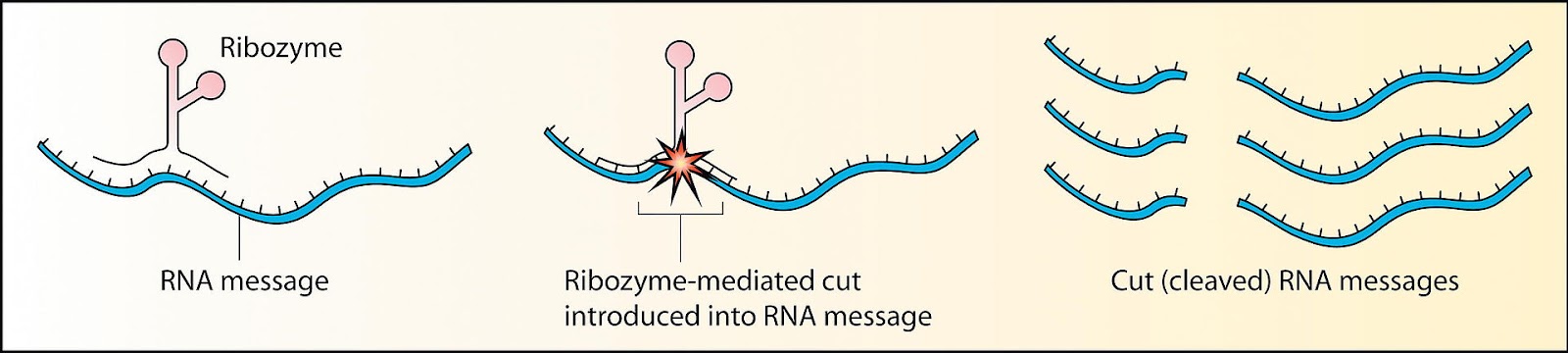
FIGURE 3. Schematic showing ribozyme cleavage of RNA. SOURCE: “RNAi Therapeutics: How Likely, How Soon?” ROBINSON 2004
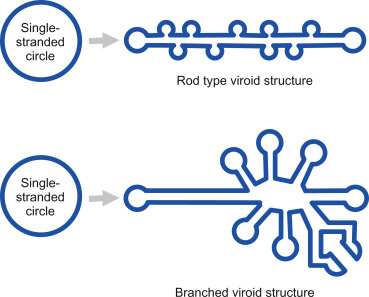
FIGURE 4. The viroid is a single-stranded piece of circular RNA that has an unusual structure due to complementary base pairing. Some form a simple rod-like structure, whereas other viroids have a complex branched structure. SOURCE: Viruses, Viroids, and Prions CLARK ET AL. 2019
The next installment of this series will delve into viroid plant diseases, with a special focus on proposed mechanisms of damage.
Read Dr. Haseltine's latest piece with
![]()
